
Imagine: Ionut Stefan
A trecut puțin mai mult decât intenționasem de la introducerea noastră despre ecuațiile diferențiale, dar sper că asta înseamnă că ai avut timp suplimentar pentru consolidarea amintirilor. Altfel, îți poți reîmprospăta memoria aici. Astăzi a venit în sfârșit momentul să abordăm neuronul virtual mult așteptat. Dar înainte să ne aruncăm în subiect, trebuie să avem o scurtă discuție organizatorică. După cum poți deja intui din lista de mai jos, de data aceasta trecem serios la treabă, așa că îți recomand insistent să citești acest articol pe bucăți. Pe de altă parte, sunt doar o voce de pe Internet și nu-ți pot spune ce să faci.
Definirea scopului
Mai întâi trebuie să înțelegem ce vrem să facem. „Construirea unui neuron virtual” sună interesant (bine, atât de interesant cât poate suna matematica), dar ne spune surprinzător de puțin despre sarcină. Trebuie să definim nivelul la care construim acest neuron. Vrem să simulăm fiecare proteină, fiecare ion și toate interacțiunile dintre ele? Poate. Recunosc că asta chiar sună destul de tare, dar am putea interpreta rezultatele? Profesorul meu de neuroștiințe computaționale obișnuia să spună: „Dacă construiești o simulare la fel de complexă ca sistemul pe care îl studiezi, acum ai două sisteme pe care nu le înțelegi.” Și, lăsând asta deoparte, am putea măcar construi o astfel de simulare acum? Ei bine, nu chiar. Așa că, în schimb, trebuie să definim trei lucruri:
- ce vrem să facem;
- ce putem și ce vrem să obținem din asta;
- ce putem să realizăm în mod realist.
Pentru astăzi, vrem să construim un model capabil să producă potențiale de acțiune, la fel ca neuronii reali (1). Vrem să folosim acest model pentru a înțelege cum produc neuronii aceste potențiale și cum sunt influențați de stimuli externi și de proprietățile canalelor ionice (2). Și putem realiza acest lucru în mod realist folosind un laptop obișnuit și propriul nostru creier (3).
Pluralitatea neuronilor virtuali
Nu există un singur mod de a simula un neuron. De fapt, există o mulțime de opțiuni. Dacă nu mă crezi, aruncă o privire aici. Alegerea unui model computațional este un act de echilibru între complexitate și eficiență. Pe de o parte, vrem ceva suficient de complex care să cuprindă ceea ce ne interesează: de exemplu, dacă vrem să știm ce se întâmplă cu un neuron când intervenim asupra canalelor sale de calciu, avem nevoie de un model care să le includă. Pe de altă parte, acest model trebuie să ruleze pe hardware-ul disponibil și ar trebui să putem înțelege într-o oarecare măsură rezultatele. Așa că, dacă ne interesează doar canalele de calciu, nu prea are sens să includem încă 300 de alte tipuri de canale ionice.
Modelul de bază
Pentru astăzi, am ales modelul Hodgkin-Huxley (HH). După cum unii dintre voi probabil știu deja, acesta este, într-un fel, piatra de temelie a modelării computaționale moderne și adesea primul „boss” pe care îl vei întâlni dacă vei urma vreodată un astfel de curs.
Deși acesta nu a fost chiar primul model computațional, modelul HH a fost revoluționar prin faptul că a fost primul cantitativ, dinamic și fundamentat biologic, și rămâne remarcabil de elegant până în ziua de azi. Desigur, acum este destul de ușor să te uiți la el și să gândești „mare scofală, știm deja cum funcționează potențialele de acțiune”. Dar, având în vedere cantitatea limitată de informații de care dispuneau Hodgkin și Huxley la acea vreme, este pur și simplu fascinant cât de bine a reprodus modelul datele empirice și ce predicții au reușit să formuleze pornind de la el.
În același timp, venind dinspre partea de biologie, am avut întotdeauna o mulțime de întrebări despre potențialele de acțiune care au rămas în mare parte fără răspuns până când am reușit să mă descurc prin jungla matematicii. De exemplu, de ce se deschid canalele de sodiu (Na+) încet la început, apoi toate deodată? De ce pragul de generare a unui potențial de acțiune are acea valoare și nu alta? De ce durează atât de mult până se deschid canalele de potasiu (K+)? Și de ce nu obținem întotdeauna un potențial de acțiune imediat după altul?
Pe măsură ce vom parcurge modelul, vom răspunde atât la aceste întrebări, cât și la altele. Dar, la fel ca în articolul anterior, vom începe cu o serie de exemple mici, inventate (codul pentru a urmări explicațiile se găsește aici) și vom avansa treptat către „bestia” principală. Sper ca aceste exemple să aducă mai multă claritate, dar dacă efectul este invers, te rog să-mi lași un comentariu. În felul acesta pot îmbunătăți acest ghid (și pe cele viitoare).
Pe parcurs, voi încerca să evidențiez atât biologia de bază, cât și ceea ce știau efectiv Hodgkin și Huxley la vremea respectivă.
De la pisici la neuroni
Abstractizarea mișcărilor unei pisici în matematică este relativ simplă. Dacă ne blocăm într-o ecuație, avem ceva concret la care să ne întoarcem. Așa că, înainte să începem cu matematica, haideți să încercăm să construim același tip de concretizare pentru neuroni și potențiale de acțiune.
Putem începe de la aceleași informații pe care le aveau la dispoziție Hodgkin și Huxley la acea vreme. Neuronii sunt înconjurați de membrane care, de obicei, blochează mișcarea ionilor. Deoarece membrana este în mod normal etanșă, putem avea concentrații diferite de ioni pe cele două părți: mai mult Na+ în exterior, mai mult K+ în interior. Deși la acea vreme încă nu știau cum sunt menținuți acești gradienți de concentrație, HH le-au recunoscut importanța.
De asemenea, au observat că, dacă plasezi un electrod în exteriorul neuronului și introduci un altul în interior, poți măsura o diferență de potențial de aproximativ -65 mV (apropo, în prezent se știe că această diferență de tensiune variază în funcție de tipul de neuron). Cu alte cuvinte, interiorul celulei este mai negativ comparativ cu exteriorul. Totuși, valoarea exactă și semnul acesteia nu contează atât de mult, cel puțin nu pentru înțelegerea principiilor generale. Ceea ce contează este că există o diferență măsurabilă și că, uneori, această diferență se modifică.
Dacă membrana ar rămâne pentru totdeauna etanșă la trecerea oricărui ion, povestea s-ar opri aici. Nu am avea niciun potențial de acțiune despre care să vorbim (și oricum nu am putea, pentru că nu ar exista inteligență, limbaj, mișcare, nimic). Dar, uneori, membrana permite ionilor să treacă prin ea. Îți poți imagina concentrațiile de ioni menționate mai devreme ca apă stocată într-un rezervor. Există mult mai mult Na+ în exteriorul neuronului decât în interior, așa că atunci când „robinetul” de Na+ (adică canalele ionice) se deschide, Na+ năvălește în neuron, asemenea apei care intră într-o cameră goală. Acest lucru se întâmplă foarte rapid și duce la o inversare temporară a semnului diferenței de tensiune: interiorul devine mai pozitiv decât exteriorul. Apoi robinetul de Na+ se închide și se deschide robinetul de K+, permițând ionilor de K+ să iasă și să readucă lucrurile la normal.
Aceste informații sunt, în mare, tot ce ne trebuie pentru modelul HH, deși sigur încă mai ai câteva întrebări.
Scurtă deviere # 1: presupus, dar nemodelat
Am menționat mai sus că Hodgkin și Huxley nu știau cum erau menținuți gradienții de Na+ și K+. Totuși, au emis ipoteza că trebuie să existe un mecanism activ care împinge Na+ în afară și aduce K+ în interiorul neuronului, contribuind astfel la menținerea gradienților de concentrație. Altfel, fiecare neuron ar avea la dispoziție doar câteva potențiale de acțiune înainte să se egalizeze concentrațiile de ioni de pe cele două părți ale membranei.
Și aveau dreptate. Ani mai târziu, s-a descoperit că există proteine încorporate în membrană, numite pompe ionice, care sunt deschise doar pe o parte a neuronului la un moment dat. Acestea acționează oarecum ca un autobuz, care permite doar Na+-ului să „se urce” atunci când pornește din interior spre exterior și K+-ului din exterior spre interior.
Scurtă deviere # 2: pozitiv, dar negativ?
Sigur nu ți-a scăpat faptul că: 1) atât Na+, cât și K+ sunt ioni pozitivi și 2) celulele, inclusiv neuronii, nu sunt încărcate electric. Așa că atunci, cum putem vorbi despre o diferență de tensiune?
Există câteva puncte-cheie aici:
- în general, cantitatea de sarcini pozitive și negative este egală atât în interiorul, cât și în exteriorul membranei, dar distribuția acestor sarcini în apropierea membranei face diferența;
- în interiorul neuronului există și proteine mari, încărcate negativ, care nu pot părăsi celula și care tind să se aglomereze în apropierea membranei;
- chiar dacă atât Na+, cât și K+ poartă fiecare o sarcină de +1, concentrația de Na+ în exteriorul celulei este mai mare decât cea de K+ în interior (aproximativ 150 mM pentru Na+ față de 100 mM pentru K+, în funcție de tipul de neuron). În plus, pompa de care am vorbit mai devreme elimină 3 ioni de Na+ pentru fiecare 2 ioni de K+ aduși înapoi, menținând astfel dezechilibrul;
- există și unele canale de K+ care rămân deschise în stare de repaus. Datorită gradientului de concentrație al K+, o parte din acești ioni ies din neuron, ceea ce înseamnă că o parte din sarcina pozitivă se scurge spre exterior, lăsând interiorul ușor mai negativ în raport cu exteriorul.
Combinația acestor factori generează diferența de tensiune măsurată de Hodgkin și Huxley.
De la neuroni la circuite
Revenind la modelul nostru de neuron, acum că avem bazele biologice, putem începe să abstractizăm. Dar, în loc să inventeze un cadru matematic complet nou pentru a descrie comportamentul neuronilor, Hodgkin și Huxley și-au dat seama că era mai simplu să adapteze ceea ce exista deja în fizica circuitelor electrice.
Toate elementele pe care le-am descris mai sus au un echivalent într-un circuit:
- deoarece membrana stochează sarcină, se comportă ca un dispozitiv numit condensator, adică un element care stochează sarcină prin acumularea acesteia pe două suprafețe apropiate, izolate una de cealaltă;
- singura modalitate prin care ionii pot trece pasiv prin membrană este prin canale ionice, care sunt de obicei închise. Cu alte cuvinte, canalele opun rezistență fluxului de ioni, așa că le putem reprezenta prin rezistori;
- am explicat și că există diferențe în concentrația ionilor între interiorul și exteriorul neuronului, iar aceste diferențe determină fluxul de ioni. Astfel, diferențele de concentrație devin sursele noastre de tensiune sau bateriile circuitului;
- și, în final, deși nu sunt incluse explicit în modelul HH, pompele ionice care refac gradienții de concentrație corespund surselor de curent din circuit, împingând ionii într-o direcție specifică pentru a menține funcționarea sistemului.

Așa cum am spus, chiar și la acea vreme exista deja multă matematică pentru lucrul cu circuite electrice. Și asta este cheia pentru a putea realiza simulările noastre.
De la măsurători la intervenții
În circuitul de mai sus, am putea măsura diferența de tensiune dintre interiorul și exteriorul membranei. De fapt, exact asta au făcut Hodgkin și Huxley la început. Au folosit axoni gigantici de la calamari și electrozi de argint pentru a măsura așa-numitul potențial de membrană sau potențial de repaus, despre care am spus că este în jur de -65 mV.
Dar măsurătorile nu sunt suficiente. Și, de una singură, starea de repaus a unui neuron și potențialul său de membrană nu sunt chiar atât de palpitante. Noi vrem acțiune… potențiale de acțiune. Acestea apar atunci când neuronii primesc stimuli sau input. O opțiune ar fi realizarea acestor măsurători in vivo, adică atunci când neuronul măsurat primește input în mod natural, fie de la alți neuroni, fie din mediul înconjurător. Dar, în situația de față, Hodgkin și Huxley și-au dorit să aibă control precis asupra inputului neuronului și să poată folosi cadrul teoretic al circuitului descris mai sus. Așa că, în schimb, au folosit un alt set de electrozi pentru a injecta curent direct în axonul unui neuron izolat.
Acum, analizând diagrama circuitului, fizica ne spune că, dacă injectăm un curent extern (pe care îl vom nota 



Cool, dar de fapt ne interesează tensiunea, nu? Până la urmă, potențialul de acțiune asta e: o schimbare în diferența de tensiune dintre interiorul și exteriorul neuronului în timp. Exact, și aici fizica ne ajută din nou: ne spune că 




Cu asta, putem rescrie 




Neuronul virtual 1 – liniar, plictisitor și instructiv
În ecuația 




Dar faptul că acest exemplu este atât de simplu nu înseamnă că nu putem extrage nicio informație din el. Observăm că, cu cât curentul input
Putem verifica și ce se întâmplă dacă pornim de la valori diferite ale lui 

Neuronul virtual 2 – până la Lună și mai departe
Acum este momentul să abordăm
Asta înseamnă că 



Observăm că tensiunea membranei crește acum mult mai repede, ajungând la valori foarte nerealiste (în practică, dacă am injecta curentul necesar pentru a atinge astfel de tensiuni, am prăji neuronul cu mult înainte de a ajunge acolo). Și dacă am varia ușor fie
Dar îți amintești cum am reprezentat canalele ionice prin rezistori? Similar cu condensatoarele, există și pentru aceste elemente o formulă care leagă curentul de tensiune: 


Iar 

Scurtă deviere # 3: acel tip pe nume Nernst
Dar stai puțin: ce legătură are concentrația ionică cu tensiunea? Și de unde provin, de fapt, potențialele de echilibru ale ionilor? Ei bine, în practică, din tabelașe drăguțe.
Dar, conceptual, trebuie să lămurim ceva, folosind Na+ ca exemplu: am spus că există mai mulți ioni de Na+ în exteriorul decât în interiorul neuronului, așadar concentrația de Na+ este mai mare pe partea exterioară a membranei. Dacă deschidem robinetul, această diferență de concentrație va împinge Na+ în interior. Dar când se oprește această împingere? Se oprește atunci când concentrația de Na+ se egalizează pe ambele părți ale membranei? Ar fi așa, dacă Na+ ar fi singurul ion prezent și nu ar exista nicio diferență de tensiune între cele două părți ale membranei.
Dar să ne imaginăm că avem și acele proteine încărcate negativ despre care am vorbit mai devreme. Asta schimbă lucrurile, deoarece chiar dacă concentrația ionilor de Na+ s-ar egaliza la un moment dat, ar exista o altă forță care l-ar atrage în interior: încărcătura negativă a proteinelor, sau gradientul electric. Deoarece aceste două forțe concurează, tensiunea efectivă la care nu mai există mișcare de Na+ este cea menționată mai sus.
Putem a calcula acest număr putem folosi încă o ecuație pe care un tip pe nume Nernst a inventat-o: ![V_{ion} = \frac{RT}{zF} \cdot ln(\frac{[ion]_{out}}{[ion]_{in}})](https://s0.wp.com/latex.php?latex=V_%7Bion%7D+%3D+%5Cfrac%7BRT%7D%7BzF%7D+%5Ccdot+ln%28%5Cfrac%7B%5Bion%5D_%7Bout%7D%7D%7B%5Bion%5D_%7Bin%7D%7D%29&bg=ffffff&fg=000&s=1&c=20201002)
![[ion]_{out}](https://s0.wp.com/latex.php?latex=%5Bion%5D_%7Bout%7D&bg=ffffff&fg=000&s=1&c=20201002)
![[ion]_{in}](https://s0.wp.com/latex.php?latex=%5Bion%5D_%7Bin%7D&bg=ffffff&fg=000&s=1&c=20201002)
Bonus: această formulă ne spune de ce influxurile bruște de K+ pot provoca decesul. Când concentrația de K+ din exteriorul neuronului crește foarte mult, potențialul de echilibru al K+-ului ajunge să fie mult mai mare decât -82 mV. La rândul său, acest lucru interferează cu generarea potențialelor de acțiune, afectând astfel comunicarea între neuroni. După ce vom avea modelul complet HH, vom putea verifica exact cum se întâmplă acest lucru.
Înapoi la neuronul virtual 2
Deocamdată, observăm că, dacă am modela doar curenții de Na+ și am presupune o conductanță constantă (în acest exemplu, 

De data aceasta, dacă schimbăm punctul nostru de plecare 

Dar ce se întâmplă dacă menținem aceeași tensiune de repaus și schimbăm conductanța 

Legat de conductanță, vreau să subliniez aici că nu este doar un concept abstract care face ca graficul să fie mai ascuțit. În viața reală, modificările conductanței canalelelor de Na+ pot avea efecte devastatoare. De exemplu, tetrodotoxina, o toxină puternică derivată din peștii pufferfish, scade efectiv conductanța Na+-ului până la zero, blocând canalele de Na+ și prevenind influxul acestuia în celulă. Acest lucru este mortal. Și în diferite tipuri de epilepsie, conductanța Na+-ului este din nou afectată: fie prea mare, fie prea mică, în funcție de tipul de epilepsie. După cum vom vedea mai târziu, modificările conductanței afectează proprietățile potențialelor de acțiune, cum ar fi forma și timpul. La nivelul întregului creier, acest lucru duce la comunicare anormală între neuroni și poate provoca simptomele observate în epilepsie.
Trecând la varierea curentului extern

Neuronul virtual 3 – alți ioni s-au alăturat petrecerii
Super, dar știm că Na+ nu acționează singur. Există cel puțin un curent de K+. Mai există și alți ioni, dar Hodgkin și Huxley au grupat tot ce ar putea acționa într-un neuron sub un așa-numit curent de „scurgere”, care este modelat ca un rezistor suplimentar.
Odată ce adăugăm curenții de K+ și de scurgere în modelul nostru (


Simularea acestui model ne permite să vedem că, la fel ca înainte, tensiunea membranei se stabilește la un punct de echilibru. Dar acest punct nu mai este egal cu tensiunea de echilibru a niciunuia dintre ioni. În schimb, se află undeva la mijloc. Această valoare de mijloc nu este nimic altceva decât media ponderată a contribuțiilor tuturor ionilor la potențialul membranei. Contribuția unui ion este dată de produsul dintre potențialul său de echilibru și conductanța sa, astfel că ecuația completă arată astfel: $latex V_{eq} = \frac{g_{Na} \cdot V_{Na} + g_K \cdot V_K + g_l \cdot V_l}{g_{Na} + g_K + g_l}&s=1$.

Am văzut mai sus că schimbarea conductanței Na+-ului atunci când doar Na+ este prezent ne permite să manipulăm cât de repede ajungem la potențialul de echilibru. Dar potențialul de echilibru în sine rămâne neschimbat. Dar acum avem mai mult de un ion, fiecare cu propria sa conductanță, și vedem în ecuația de mai sus că potențialul de echilibru al membranei ia în considerare și conductanțele. Așadar, ce se întâmplă dacă schimbăm fiecare conductanță ionicǎ individual?
Ar trebui să putem deduce acest lucru din ecuație, dar îl vom verifica cu rezultatele simulării de mai jos. Linia albastră reprezintă cazul nostru original de mai sus. Deoarece potențialul de echilibru al K+-ului este mai negativ decât potențialul nostru de repaus original 

În principiu, am putea să experimentăm și cu conductanța de scurgere 
Modelul complet – toată lumea primește o funcție
Am adăugat deja destule detalii modelului nostru, dar mai este încă puțin de lucru. Până acum, avem o simulare a tensiunii membranei care include multiple canale ionice. Acest model este capabil să se stabilizeze la un punct de echilibru, potențialul de repaus, dar încă nu produce vârfuri (spikes). Așadar, haideți să remediem asta. Un mic avertisment: partea următoare este cea mai dificilă (știu! Ca și cum romanul de până acum ar fi fost așa ușor!), așa că mergi încet, fă pauze frecvent și nu îți face griji dacă va fi nevoie de câteva lecturi pentru a înțelege.
Punct cheie # 1: conductanțele sunt dependente de tensiune
Să facem acum puntea între biologie și matematică: am spus că atunci când conductanța Na+-ului crește (adică se deschid canalele de Na+), tensiunea membranei crește și ea. Dar știm și din experimente că atunci când tensiunea membranei crește, canalele de K+ se deschid. Cu alte cuvinte, conductanța K+-ului crește. În termeni matematici, asta sugerează dependența conductanțelor (pentru Na+ și K+) de tensiune.
Punct cheie # 2: există o conductanță maximă
Imaginează-ți că toate canalele de Na+ sunt deschise. Chiar și atunci, există totuși o limită la cât de mult Na+ poate trece prin membrană la fiecare pas de timp, pentru că ionii trebuie să aștepte pe rând pentru a trece prin canale, la fel cum mașinile trebuie să aștepte pentru a trece printr-un tunel aglomerat. Asta înseamnă că există o valoare maximă a conductanței, pe care o putem numi 


Punct cheie # 3: putem lucra direct cu proporțiile canalelor deschise
Dar ce se întâmplă dacă doar 50% din canale sunt deschise? Ei bine, limita ar fi jumătate din valoarea maximă: 
Punând totul împreună
În primul rând, având în vedere dependența conductanțelor de tensiune și că tensiunea membranei se schimbă în timp, avem, de fapt, conductanțe dependente de tensiune și timp. Este important de menționat că acest lucru se aplică doar pentru Na+ și K+; conductanța de scurgere este fixă.
În al doilea rând, lucrăm cu proporția de canale deschise, nu cu conductanțele direct. Haideți să facem o pauză și să ne gândim la ce vrem să modelăm. Practic, vrem un fel de mecanism de tip push-pull, astfel încât atunci când tensiunea crește, proporția de canale de Na+ deschise să crească, iar când tensiunea scade, proporția de canale închise să crească. Și la fel pentru K+.
Să începem cu K+. Putem nota proporția de canale de K+ deschise cu n. Proporția de canale închise va fi pur și simplu 1 – n (totalul minus câte sunt deschise). Deoarece suntem interesați de cum evoluează acest lucru în timp, trebuie să aducem înapoi vechiul nostru prieten, ecuația diferențială, în acest caz 


- cele două funcții A și B acționează ca factori scalare pentru proporția de canale închise, respectiv deschise. A controlează cât de repede se deschid canalele închise, iar B controlează cât de repede se închid canalele deschise.
- ecuația de mai sus nu este suficientă. A și B sunt funcții dependente de tensiune și trebuie alese astfel încât, atunci când tensiunea crește, A să crească și B să scadă, iar invers atunci când tensiunea scade.
Dar cum le alegem? Ei bine, ecuația de mai sus este o ecuație diferențială de ordinul întâi și are o soluție cunoscută. Fără a intra în detalii matematice, Hodgkin și Huxley au folosit această soluție împreună cu măsurători experimentale ale curentului de K+ pentru a deriva formule specifice pentru 



(Notă: convenția semnului) Un lucru de remarcat mai sus este că folosim atât 



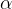

Pentru Na+, HH au modelat activarea canalelor de Na+ într-un mod similar, doar că au denumit proporția de canale activate m. Din nou, pentru completitudine, ecuațiile respective au fost 

Acum avem aproape întregul model HH funcțional, dar mai sunt doar câteva ajustări minore. Deoarece Hodgkin și Huxley și-au ajustat modelul la date experimentale, au observat două detalii interesante:
- modelul se potrivea mai bine datelor experimentale când variabila n era ridicată la puterea a 4-a și când m era ridicat la puterea a 3-a. La vremea respectivă, nu știau de ce se întâmpla acest lucru, dar între timp am aflat că pentru ionul de K+, canalul este format din 4 subunități, fiecare dintre care trebuie activată pentru acesta să permită trecerea K+-ului. În acest caz, poți considera n ca fiind proporția de canale în care subunitatea 1 este activată (sau probabilitatea ca această subunitate să fie activată). Proporția de canale în care 2 subunități sunt activate este
, și așa mai departe. Similar, canalele de Na+ au 3 domenii de activare care trebuie deschise pentru ca Na+ să treacă prin canal;
- când tensiunea membranei era menținută constantă la o valoare mare, K+ continua să iasă din celulă până când tensiunea revenea la valoarea normală. Dar pentru Na+, Hodgkin și Huxley au observat un comportament diferit: Na+ a intrat în celulă la început, apoi s-a oprit. Curentul de Na+ a scăzut brusc și, indiferent de cât timp tensiunea a fost menținută ridicată, curentul de Na+ nu a mai crescut. Pentru a modela acest comportament, au introdus o a doua variabilă pentru Na+, numită h, pe care au folosit-o pentru a modela proporția de canale de Na+ inactivate. Aceasta a necesitat aceleași funcții
și
, cu
și
. Din nou, în zilele noastre știm că Na+ are un domeniu de inactivare care blochează rapid canalele de Na+ la tensiuni mari și le deblochează doar atunci când tensiunea scade din nou. Acesta este și motivul pentru care potențialele de acțiune nu se pot răspândi înapoi de la locul de unde au venit.
Și asta e tot, acum avem un model HH complet. Punând totul laolalt, arată cam așa:


Este important de menționat că, de unul singur, modelul nu face de fapt nimic. Dacă inputul extern este zero și inițializăm modelul de la o tensiune de membrană sub un anumit prag (în acest caz, 

Dacă inițializăm modelul deasupra unui anumit prag (de exemplu,

Pentru a obține mai multe vârfuri, trebuie să-l alimentăm cu curent extern. Până acum, am folosit curent constant și vom rămâne cu acesta și astăzi (în partea următoare, vom încerca și curenți variabili în timp). Pentru un curent suficient de mare, vedem că modelul declanșează un potențial de acțiune după altul. Poți încerca și tu să vezi ce se întâmplă pentru valori diferite ale

În final, putem inspecta variabilele noastre de control m, h și n, pentru a vedea cum evoluează în timp. În graficul de mai jos, vezi că variabila de activare a canalelor Na+, m (în albastru), crește foarte repede – canalele Na+ se deschid rapid; dar scade la fel de repede – ele se închid rapid. Variabila de inactivare a Na+, h, scade rapid în timpul vârfului – canalele Na+ sunt blocate și nu pot să se deschidă din nou pentru o perioadă de timp. Între timp, variabila de activare a K+-ului, n, crește, rămânând puțin în urma lui m – canalele K+ se deschid mai lent și tensiunea membranei scade înapoi.

Mai trăiește cineva?
Nu știu despre tine, dar eu am obosit. Vestea bună e că acum avem un model HH funcțional. Altă veste bună e că putem face multe lucruri cu el, dar din păcate, asta necesită explicații suplimentare, și cred că tuturor ne-ar prinde bine o pauză. Așa că ne vedem la următoarea parte. Până atunci, nu ezita să te joci cu parametrii modelului.
P.S.: Dacă cineva știe o soluție mai bună pentru afișarea ecuațiilor LaTeX în WordPress, să mă anunțe, vă rog. Metoda actuală îmi dă dureri de suflet.
Cum ți s-a părut această postare? Scrie-ne în comentariile de mai jos. Și dacă vrei să ne susții, poți distribui articolul, ne poți cumpăra o cafea aici sau chiar ambele.
Abonează-te la fluxul RSS aici.
Ar putea să-ți placă și:
Referințe (în engleză)
Goaillard, J.-M., & Marder, E. (2021). Ion Channel Degeneracy, Variability, and Covariation in Neuron and Circuit Resilience. Annual Review of Neuroscience, 44(1), 335–357. https://doi.org/10.1146/annurev-neuro-092920-121538
Hodgkin, A. L., Huxley, A. F., & Katz, B. (1952). Measurement of current‐voltage relations in the membrane of the giant axon of Loligo. The Journal of Physiology, 116(4), 424–448. Portico. https://doi.org/10.1113/jphysiol.1952.sp004716
Hodgkin, A. L., & Huxley, A. F. (1952). A quantitative description of membrane current and its application to conduction and excitation in nerve. The Journal of Physiology, 117(4), 500–544. Portico. https://doi.org/10.1113/jphysiol.1952.sp004764


Lasă un răspuns